Robert A. Freitas Jr., “Extraterrestrial Zoology,” Analog Science Fiction/Science Fact, Vol. 101, 20 July 1981, pp 53-67
URL: http://www.xenology.info/Papers/ETZoology1981.htm
This paper contains material originally drawn from the book Xenology (1979) by Robert A. Freitas Jr.
“Two large dark-coloured eyes were regarding me steadfastly.
The mass that framed them, the head of the thing, it was rounded, and had, one
might say, a face. There was a mouth under the eyes, the lipless brim of which
quivered and panted, and dropped saliva. The whole creature heaved and pulsated
convulsively. A lank tentacular appendage gripped the edge of the cylinder,
another swayed in the air. ... There was something fungoid in the oily brown
skin, something in the clumsy deliberation of the tedious movements unspeakably
nasty.”
– H.G. Wells, The War of
the Worlds (1898)
Pretty disgusting, huh? The classic tales of science fiction are full of Bug-Eyed Monsters (or BEMs as they are affectionately termed by cognoscenti) which invade planets, threaten towns. attack rocket ships, and carry off shapely human females. Hollywood producers apparently are convinced most extraterrestrial (ET) beings fall in one of four zoological categories: (1) Human or humanoid, (2) oversized animals, (3) amorphous blobs and pods, and (4) formless energy beings.
Can’t we do any better than this?
Quite! In fact. anyone with access to a good library can walk in and read all about the biology of one of the most fascinating, richly populated worlds anywhere in the Milky Way: Earth! We inhabit a queer planet with many strange settings and fabulous living creatures, altogether an excellent example of what extraterrestrial life may be all about. To a team of Interstellar Zoologists, researching sentient terrestrial mammals out here in the galactic boondocks, our world is as rare a planetary zoo as any in the Milky Way.
Xenobiologists have formulated a simple rule called the Assumption of Mediocrity, which says, in essence, that Earth should be regarded as “typically exotic.” The unusual solutions devised by evolution on this planet to cope with the problem of survival will find their parallels, though not necessarily their duplicates, among the living species of other worlds. As biologist Allen Broms once remarked, “life elsewhere is likely to consist of odd combinations of familiar bits.”
Life as we know it is based on cells: small, neat packages of living protoplasm containing all of the biological machinery necessary for survival. Human body cells average a few microns in size. (One micron is a millionth of a meter, about a hundredth of the thickness of the page these words are printed on.) The smallest living thing on Earth capable of independent metabolic activity is the PPLO, or “pleuropneumonia-like organism,” which measures 0.1 microns. Microbiologists estimate that the smallest cell that could, in theory, exist would measure about 0.04 microns in diameter. It is amusing to speculate that the alien analogue to a human being, constructed in the same form but using these miniature cells, would weigh a mere 50 milligrams and stand only 5 millimeters tall – hardly the thickness of a pencil. Whether creatures so small could retain a human-level intelligence is anyone’s guess.
Fairly large extraterrestrial lifeforms might well exhibit acellular physiology, or be unicellular. For example, at one stage in their life history, slime molds are tiny one-celled flagellates capable of individual multiplication by simple fission. In the later “plasmodium” stage of development, large clumps of these creatures fuse together and their cell walls dissolve away to produce an amorphous acellular mass of living protoplasm which can grown as large as 25 centimeters or more. Further, the largest known single living cell was the egg of the now-extinct half-ton elephant bird or “roc bird” (Aepyornis maximus). This egg measured about a third of a meter across and weighed 15 kilograms.
The number and kinds of organs in alien creatures may also be highly variable. For example, earthly squids have two different kinds of hearts – one for venous and a separate one for arterial blood – and the common earthworm (Pheretima) has a dozen hearts. Two extinct dinosaur species, Brontosaurus and Diplodocus, had two brains, one in the head and an even larger hunk of neural tissue in the hip region. (The volume of this “sacral enlargement” in Stegosaurus, another fossil animal of grand proportions, was perhaps twenty times larger than the brain in the cranial cavity! And the entire body of an insect is its “lung” – oxygen is carried directly to cells by an intricate network of tracheae or microtubules permeating the entire organism.
Sometimes, organs combine several functions in one – such as the human mouth. ETs need not have the same combinations as we. They may have identical or separate organs for eating, drinking, excreting, breathing, and speaking. The dolphin, for instance. eats through its mouth, breathes through its blowhole, and “speaks” through its “ears.” The land snail’s lung opens into a passageway other than its food canal, and sea cucumbers breathe through their rectums (called “anal respiration”). The cloacae of frogs and many other animals is a single organ which combines excretory and reproduction functions. Brachiopods can only vomit excrement from their “blind intestine” (a kind of alimentary cul-de-sac), and the members of phylum Nematomorpha (long worms) eat solely by direct absorption of nutrients through the skin – for they have no mouths.
How big can ETs be? To answer the question we need to understand something called the Square-Cube Law. This universal geometrical principle, first recognized by Galileo more than three centuries ago, holds that volume always increases faster than surface area as size increases. A solid cubical box whose edge is doubled increases in surface area by a factor of two squared (2x2), or four; whereas volume, hence mass, increases by two cubed (2x2x2), or eight.
It’s easy to apply this to biology. Picture a bony extraterrestrial herbivore placidly grazing in some alien meadow. Suddenly we double its size all over. The animal’s leg bones, now twice as thick, have quadrupled in cross-sectional area; but the creature weighs eight times as much so its bones must sustain double the pressure. It may collapse under normal exertion unless it grows proportionally stouter limbs to handle the added physical stress.
All parts of an animal must be reengineered when size increases. Like bone, muscle strength is determined by cross-sectional area. Humanoids twice as large need quadruply thick biceps: otherwise they’d be pulling eight times the mass with only four times the force. Lungs, kidneys, intestines and other blood filtering organs function according to surface area, so must either increase in mass or become more convoluted at larger body sizes.
The horror movies about giant insects ravaging the countryside are really quite impossible, even on low-gravity worlds. A bug as large as a house would weigh a billion times more than its flea-sized Earthly cousins. Its thin spindly legs would be called upon to sustain stresses thousands of times greater. To walk at all the overgrown arthropod needs muscles proportionally thousands of times thicker; unfortunately, vital tissues already fill the hollow skeleton of the tiny original. It did not collapse under its own weight or was not immobilized by the feebleness of its muscles, an overgrown insect would starve to death because its stomach would be a thousandfold too small to absorb enough food; or it would suffocate because its tracheae could carry only a thousandth as much air as needed.
Sea creatures are free of gravity at neutral buoyancy, but still they’re dogged by the Square-Cube Law. Bodies in motion like to continue in motion – extraterrestrial leviathans larger than whales would experience serious steering, turning and braking difficulties because of their relatively great mass compared to the area of their control surfaces. Cornering too fast might cause stresses in excess of the tensile strength of biological materials and the behemoth would literally snap in two. These problems are familiar to pilots of modem supertankers, huge ships requiring kilometers to turn or stop.
The respected zoologist D’Arcy Wentworth Thompson once speculated about the effects of gravity on evolution. “Were the force of gravity to be doubled,” Thompson declared, “our bipedal form would be a failure, and the majority of terrestrial animals would resemble short-legged saurians, or else serpents. Birds and insects would suffer likewise, though with some compensation in the increased density of the air. On the other hand, if gravity were halved, we should get a lighter, slenderer, more active type, needing less energy, less heat, less heart, less lungs, less blood. Gravity not only controls the actions but also influences the forms of all save the least of organisms.”
It is true that the maximum weight of living species cannot exceed the crushing strength of bony material. But animals are not designed to stand still – if they were, human legs could be a few millimeters thick. Instead they must bear up under the peak pressures and accelerations encountered during normal running, jumping, and other strenuous survival activities. A horse at rest seems greatly overbuilt; on the racetrack where it may pull to a halt in a second or less, near the breaking point of its bones, the design limits are more fully exploited.
Clearly there are other factors at work besides gravitational loading in fixing maximum size – predator/prey relationships, running speeds, food requirements, oxygen levels, ecological constraints, and so forth. Still we can estimate how gravity might influence evolution, based on Earth’s biological history. The largest land creature alive today is the African elephant, weighing an impressive 6600 kilograms. Tyrannosaurus rex, one of the largest land carnivores, was at least 8000 kg. The Baluchitherium, the largest extinct land mammal, was built like a hornless rhinoceros, and carried a bulk of more than 22,000 kg. The largest land animal ever may have been Brachiosaurus, of which some specimens may have weighed 111,000 kg. but we’ll ignore this majestic brute because he probably had to spend lots of time sitting in swamps resting his tired bulk. We may conservatively guess that the heaviest exclusively land-dwelling creature plausible on a 1-gee planet is around 22,000 kg.
How massive will alien animals be? Simulations of model solar systems by Dr. Stephen H. Dole of the RAND Corporation and others suggest that terrestrial rocky worlds with atmospheres suitable for life should have surface gravities between about 0.2 and 2.0 Earth-gees. Now, if gravity doubles, bone stress won’t increase if a creature’s height is halved while other dimensions remain the same. If maximum height is inversely proportional to gravity, then maximum volume (hence mass) goes inversely as gravity cubed. By this measure the heaviest animal on a 2-gee world is about 2800 kg, while on a 0.2-gee planet (like Saturn’s moon Titan) the most massive beast could conceivably reach nearly three million kilograms – though I’d hate to try to keep it fed! So animals like walruses, small elephants, even 70 kg humanoids are quite possible even on the heaviest of all reasonable Earthlike worlds. No need for “powerfully built, squat creatures, perhaps rather like an armoured pancake on multiple legs ... limited to slow, creeping motions across the surface.”
Of course, gravity will affect design. In any given mass category high-gee animals should have shorter, stockier bones than those evolving in low-gee environments. To provide proper support, bone cross-section must increase directly with weight. Weight is the product of mass and gravity, so bone diameter must be proportional to the square root of gravity.
Let’s apply this to man. The typical human femur, the most perfectly cylindrical and largest single bone in our bodies, measures 3.5 centimeters in diameter. Using the above square-root relation, we find that the thigh-bone should increase to 4.9 cm on a two-gee world or shrink to 1.6 cm on a 0.2-gee planet for identical support of a 70 kg human body mass. Experiments have confirmed that animals reared in high gravity grow thicker bones, stronger hearts, and lose fat, but alien creatures will not appear wildly over- or underbuilt as compared with Earth life of equal mass.
Boneless lifeforms in the sea can grow to enormous sizes. There are other advantages to life without a rigid frame we can hardly appreciate. For instance, an octopus, often called the supreme escape artist, can stretch itself incredibly thin, passing rubberlike through small holes or narrow crevasses and sliding confidently across desktops and the decks of ships.
But a creature of land is a denizen of gravity. Surface life must evolve some means of physical support or be reduced to a groveling mass on the ground. On Earth the most common frameworks are the exoskeleton and the endoskeleton. The former, typified by insects and crustaceans, is a hollow bony tube packed with the creature’s viscera. The latter, which all vertebrates have, is a central spine from which vital organs hang like coats on a hat rack. Exoskeletons are bony material surrounding gut; endoskeletons are bone surrounded by gut.
Which design is better? Bioengineers point out that a tubular column always has greater strength than a solid beam of the same mass. Tubes give twice the resistance to bending and many times the opposition to buckling. Mechanical advantages are best exploited by exoskeletons because of the greater bony surface area to which muscles may be attached.
So why be vertebrate? The answer is that we’ve considered only static strength. Large endoskeletons outperform exoskeletons under dynamic impact loading – like falling out of trees – which is why the largest of all animal species have worn their bones on the inside. Massive alien insectoids are not impossible, just less likely. Falling impacts shouldn’t be as severe on low gravity planets, and large active arthropods might survive in a rich oxygen atmosphere. The greatest carapaced creatures on Earth have ranged in size from a tenth of a meter for the South American tarantula on land up to several meters for certain fossil marine arthropods.
ETs have other choices open to them. One of the most popular alternatives among xenobiologists is called the “basket skeleton” found on this planet in marine echinoderms (sea cucumbers. starfish, sea urchins) and the cormorant (a seabird of the pelican family). Physical stress passes through the body along a kind of bony trellis, an unusual internal arrangement which one wag has facetiously termed “bowels in a birdcage.”
Another possibility is the double spine or multiple endoskeleton. On Earth flatworms and other free-living turbellarians have twin neural channels running the length of their bodies. Alien “ladder skeletons” might improve postural stability and provide greater strength on high-gravity worlds, though turning or twisting motions of the trunk might be restricted even if the multiple support posts are jointed or segmented.
A third alternative is the “hydrostatic skeleton,” surprisingly common on Earth. Animal bodies are kept stiff by pressurized fluid trapped in a sack of tough skin. Mostly only small earthworms and nematodes have this support, but massive sea creatures such as sharks compress their innards to help negotiate sharp turns and even man uses the contents of his abdomen as a hydrostatic skeleton. Large aliens might evolve a liquid skeleton inside taut, fiber-strengthened tubes with extensive reinforcing musculature – purely hydrostatic caterpillars, for example, have about 4000 individual muscles as compared to less than 700 for a human being.
Nature often uses the same solution to a given problem encountered by many independently evolved species. Perhaps one of the most striking instances of this “convergent evolution” is the “camera eye’’ invented separately by at least five major terrestrial animal phyla (chordates, mollusks, annelids, coelenterates and protists). Each have radically different developmental histories. Naturally there are a few discrepancies – for example, light-sensitive cells in molluscan eyeballs point towards the light, the opposite of vertebrates. But the adjustable lens. retina, pigments, focusing muscles, iris diaphragm, transparent cornea and eyelids all are immediately recognizable. Nature is perhaps trying to tell us something: The camera eye is ubiquitous because it’s simply the best design for the job, on this or any other world.
The next most successful – indeed more so if you just count species – is the compound eye of insects and crustaceans. Each organ looks like a small multifaceted jewel, actually a tiny bundle of optical tubes that direct light onto a large matrix of individual photosensitive spots on the retina. The image forms a composite mosaic of thousands of little light-dots. (Dragonfly eyes have more than 28,000 facets and can discern motion up to twelve meters away.) The compound eye, however, has such poor resolving power that an insect poring over this page of print would be quite unable to make out the individual letters, so large ETs will find the system unattractive. It seems best for smaller creatures – if a flea had a spherical lens eyeball like that of humans, the pupil would be so minute that diffraction effects would utterly ruin the image.
Other visual techniques of limited importance on Earth may be emphasized on other planets. For instance, alien species may have “pinhole camera” eyes like the chambered nautilus, a beautifully simple system consisting of an open optical pit without lenses, exceptionally useful in water. In the “scanning eye” of the snail, light penetrates a simple crystalline lens and is scanned by a single retinal nerve sensor moving across the visual field, slowly building up an image of the environment. The principle of the optical reflector telescope has never been developed for direct imaging on this world, though many species use a biological mirror assembly to increase camera eye sensitivity (the tapetum of the common tabby cat) or to attract prey using deep-sea “searchlights” in conjunction with bioluminescence (the retractable reflectors of the luminous squid).
How many eyes are best? Nature usually economizes, so a single receptor organ is good enough for nondirectional sensing. Most large organisms have but one organ of smell and one of taste. On the other hand, directional senses can make good use of the benefits of stereo. Triangulation and depth perception require at least two physically separated receptors, and there seems little to be gained by going to more than a single pair. As astronomer Carl Sagan once pointed out, “Three eyes represent not nearly the same improvement over-two that two represent over one.”
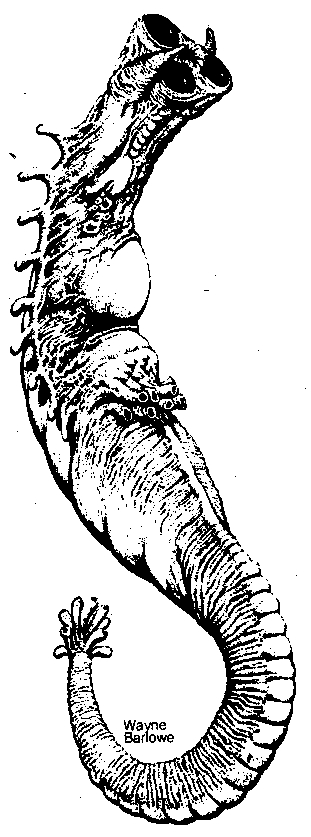
Figure 1. A Three-Eyed Alien Lifeform (courtesy of Wayne Barlowe)
Nevertheless a few animal species do have more than one pair of imaging eyes. Zoologist Norman J. Berrill of McGill University in Montreal describes the dinnertime antics of the spider, which has four pairs of eyes: “The rear pair serve to watch behind for either food or danger. The other three pairs work together but in succession. If something comes within the range of vision of one of the outermost pair, the head turns until the object is brought into the field of the two pairs of eyes in the middle, and the spider then advances. When the object is brought into focus of the forward pair, the spider jumps to attack.” The ultimate limit is probably reached by the scallop, whose literally hundreds of tiny, beautifully constructed nonimaging “eyes” are spread around the circumference of its mantle like running lights on an ocean liner.
What about eyes on stalks? Most xenobiologists regard this as a rather unlikely adaptation for thinking animals. Eyestalks require a hydraulic support system inefficient except in small animals. Eyes are vital senses for large organisms, yet stalks could be lopped off by predators with a single stroke of claw or pincer, permanently depriving the owner of sight. Periscoping eyes unprotected by bone are also more prone to common injury – in an accident, stalks could be bumped, slammed or squashed all too easily.
Vision, of course, is simply the detection of one narrow set of wavelengths of light within the entire electromagnetic spectrum. One alternative to “visual” sight is infrared (IR) vision, or seeing with heat waves. The rattlesnake is quite good at this – the creature has two imaging eyeballs operating in the visible, and two conical pits on either side of the head which permit binocular IR sensing of temperature differences as little as 0.002 °C. The theory of optics predicts that alien infrared eyeballs with resolution close to that of the human eye could have apertures as small as 4 centimeters at 93,000 Angstroms (the peak wavelength of black body radiation emitted by a warm human body). This compares well with the size of the eye of the Indian elephant (4.1 cm), the horse (5 cm), the blue whale (14.5 cm), and the largest cephalopods (up to 37 cm).
Radio vision is another possibility, although there are two major evolutionary problems with this. First, it is difficult (though not impossible) to imagine planetary surface conditions in which the illumination in the radio band is equal to or greater than the brightness in the visible, thus giving radio vision the competitive edge. Second, radio sensors would have to be on the order of 10-1000 meters wide to achieve human-eye acuity, though this resolution may not be absolutely necessary.
Assuming life evolves primarily on planetary surfaces and under air, other forms of vision – very low frequency, ultraviolet, and x-ray – are unlikely because these wavelengths are strongly absorbed during the passage through atmosphere or ocean. Static electric field sensing has been documented in numerous species, notably sharks and electric fishes, and sensitivity to magnetic fields has been found in snails, pigeons; dolphins, bees. and many other animals. The acoustical, tactile, and chemical spectra of sensation have also been well exploited by life on Earth.
One possible extraterrestrial sense often overlooked is the ability to detect radioactivity. On a world with highly concentrated radionuclide ores near the surface, or on a planet in the throes of a global nuclear holocaust, biological Geiger counters would give warning to steer clear of large tracts of radiation hazards. The “radioactive sense” was once artificially bestowed on a small group of laboratory animals by wiring portable Geiger counters directly to the fear center of feline brains. When confronted with a pile of radioactive materials in one comer of their cages, each cat shied away.
The key to alien senses is survival – any environmental information that would permit an animal to better compete for the limited resources available is a valid candidate for sensing. For example, we could imagine a sophisticated meteorological sensorium evolving on a world cursed with highly volatile, perpetually inclement weather. Humidity and barometric sensors would be essential, as would anemometers to calibrate wind velocity. The ability to sense changes in atmospheric composition, such as the carbon dioxide detectors possessed by honeybees and fire ants, would be useful. Atmospheric turbidity, closely related to developing weather patterns, greatly influences the degree of skylight polarization – sensors responsive to the intensity and distribution of polarized light might permit their owner to seek shelter from the elements before disaster struck. The seeming ability of many animals to sense an earthquake or tornado before it arrives may relate to their perception of very low frequency infrasonics or minute electrical field variations immediately preceding the event. And the allegation that elephants can sense water located a meter or so beneath the surface of apparently dry riverbeds is unproven scientifically, yet the fact remains that such biological dowsers would be tar more likely to survive on a drought-stricken planet.
On strictly mechanical grounds, three points are needed geometrically to define a surface plane – two points make only a line. ETs trying to stand up on just one or two levers will promptly fall flat on their faces. We bipedal humans manage to remain erect only because our large feet provide additional points of contact with the ground, but without toes or feet a minimum of three legs is necessary.
Are tripedal aliens possible? Traditional biologists say no. A walking three-legger must lift at least one limb off the ground, at which instant it loses its planar support base, a situation statically unstable and dynamically precarious. Four legs seem better from an engineering point of view, as the creature can remain balanced when a leg is in motion. Ancestral fishes only have fins in pairs, so mustn’t all limbs evolve in pairs as well?
Xenobiologists remain unconvinced. Most running bipeds and quadrupeds keep two or fewer limbs on the ground during locomotion, so three-point dynamic stability is probably unnecessary. Land life need not always evolve from pair-finned fishes – descendants of, say, a starfish might be odd-leggers. Most persuasive, however, is the simple fact that tripeds exist on Earth! The extinct Tyrannosaurus rex and a few large contemporary creatures such as the kangaroo run bipedally but stand tripedally. The tails of these animals are as strong and thick as the forelegs and are regularly used for postural support. Indeed, when kangaroos fight, they rear up on their tails, freeing both legs to deliver crushing kicks to opponents.
More legs than four are plausible even for massive, intelligent animals. Odd appendages are often used for highly specialized purposes, as witness the prehensile tail of monkeys and the dexterous trunk of elephants. The key to higher multipedia is neural control. The nervous circuitry for an extra limb is far less than that required to add, say, another eye. Muscles need thousands of new neurons, but eyeballs need millions. About one-third of the mammalian brain is committed to sensory functions, whereas only a small slice handles motor control, ETs are much more likely to have extra arms than extra eyes or ears.
Dr. Bonnie Dalzell, a writer-paleontologist who helped Larry Niven work out some of his fictional aliens, insists that vertebrates on Earth have four limbs solely because of the common descent from fishes adapted to free-swimming conditions in large open oceans. These fish needed only two independent sets of diving planes to make a go of it in the sea. Perhaps if we evolved instead from Euthacanthus, a Devonian Period fish boasting no fewer than seven pairs of fins, we might be hexapodal or more-podal today ourselves.
Dr. Dalzell expects to find intelligent six-leggers on worlds with small, shallow oceans. There, bottom-dwelling fishes would become the predominant coastal and freshwater lifeforms early in evolutionary history. If the planet has a very seasonal climate, perhaps accompanied by large-scale periodic evaporations of lakes and seas, few fish species could evolve into good swimmers as on Earth. Marine creatures with many pairs of fins would have the advantage, ultimately inheriting the land and producing a rich ecology of multipodal animal life.
There are many advantages to six-legged living. On high-gravity worlds hexapedia is a good way to distribute mechanical stresses and help reduce the danger of bone breakage. Injury or loss of a limb is more catastrophic for four-leggers than for six-leggers (who have “spares”). Hexapods also have better balance since, unlike quadrupeds, they can keep a stable support tripod on the ground even when running at high speeds. And it shouldn’t be too hard to coordinate all those legs. Says Dalzell: “Earthly insects with three pairs of legs are hardly noted for their well-developed mental powers, but most of them walk just fine.”
Figure 2. Hexapodal Alien Animal (courtesy of Wayne Barlowe)

Of course, legs are not the only game in town. The potential of rotary motion (to pick one possibility of many) cries out for fulfillment. A few years ago biologists made the amazing discovery that the tails of tiny bacteria are driven by minute ionic motors complete with rotors, stators, bushings and freely-rotating drive shafts spinning up to 60 cycles per second. The rapid back-and-forth wiggling of flagella we see under the microscope is actually a complicated helical twisting movement more akin to a propeller screw than to a simple fishy undulation. This finding contradicts the long-standing dictum that living organisms may not contain detached, self-rotating parts.
Rotary motion may be possible for large animals too. Picture a small Earthlike world with little tectonic activity and broad, flat continental shelves flooded to a depth of five or ten meters during global warm spells. A creature not unlike the molluscan cuttlefish Sepia hovers near the bottom, stalking small fish, shrimps, and crabs, sometimes jetting about by expelling water rapidly from several exit portals like many other cephalopods. Occasionally sand particles jam in a portal, causing irritation. The animal responds by encasing them in a perfectly smooth spherical pearl, much like those of the modem oyster.
Millions of years later an Ice Age arrives. The retreating shoreline leaves behind vast tracts of smooth hard continental shelf. Forced into ever more turbid, colder, shallower waters, we might imagine our cuttlefish eventually abandoning the sea for land, evolving into a “caster creature.” Its jet ports now permanently plugged by large pearly structures almost from birth, these animals might develop the ability to roll along the graded continental raceways. Speed is controlled by internal sphincters aided by heat sensors for guided braking on gentle downhill stretches and a “low-gear” muscular assist for steep climbs. Tentacle arms like ski poles provide additional stability on fast runs along the coastline.
How big could flying ETs evolve? On Earth the albatross is pretty close to the maximum. This 10 kilogram bird reaches wingspans up to four meters and needs a lengthy runway to achieve takeoff speed of 20 kph. This minimum velocity is called the “stall speed” and is partly determined by air density. Venusian pigeons could remain airborne at speeds ten times slower than their Earthly cousins, whereas Martian birds of similar size and shape would have to fly ten times faster to stay aloft.
The main factor fixing avian size is atmospheric pressure, not gravity as some erroneously believe. On high-pressure worlds, alien bird creatures can have surprisingly small wings and large masses. An extraterrestrial with the mass of a man could fly with the wings of an albatross in air just five times thicker than Earth’s, and a Venusian albatross could make do with stubby wings smaller than the page on which these words are printed.
Planetary surface gravity has less effect on size in part because it varies far less than air density from world to world. For the same ease of flight a pigeon on a 2-gee planet with Earthlike air must increase total wing area by only 75 percent; on a bantam-weight 0.2-gee world, wing surface may decrease 75 percent. Gravity also influences stall speed. An albatross on a 2-gee planet needs a 40-percent runway extension; on a 0.2-gee world it can get by with 55 percent less. Massive extraterrestrial avians are more likely on puny planets with dense atmospheres.
How many wings are best? Most common among terrestrial species is a single pair which generate lift by actively beating the air something like the blades of a helicopter rotor. Less common is the “airplane” system, with one pair producing passive lift (like the wings of an airplane) and a second pair taking the more active role (like propellers). Adding yet more wings would serve no useful purpose, hence are unlikely to evolve. Only a very few insect species on Earth retain vestigial traces of an ancestral third wing pair, and these are degenerate and useless for flight.
Alien air travelers may have no wings at all! There are many alternatives that have never been fully exploited by evolution on this planet. Consider, for example, the principles of the rocket, the glider, and the balloon.
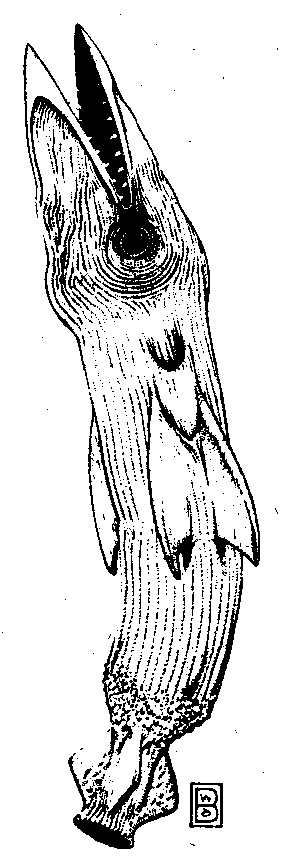
A high-gravity world with abundant seas and a warm, thick oxygen atmosphere might produce a “rocket fish” predator, patrolling the coastal shallows and preying on bird-sized torpid insect life thickly swarming high up. Much like the toy plastic projectiles that shoot the length of a playing field when fully charged with water and compressed air, the rocket fish bolts from the sea skyward and mouthes its dinner on the fly. Such an animal must have a sturdy posterior pressure canister that can be discharged rapidly through a rigid bony nozzle, rechargeable in minutes using powerful sphincter muscles, internal gas generation, or osmosis. Earthly precedents include the jet propulsion of squids and octopuses, the pressurized chemical sprays of warrior termites, and the boiling liquid jet of the bombardier beetle.
Figure 3. Illustration of the Rocket Fish (courtesy of Wayne Barlowe)
A lightweight planet with high winds might be ideal for the evolution of sentient “parachute beasts,” large aerial aliens able to navigate the airways of their world by manipulating sturdy chutes or simple gliding surfaces. Vultures can sail for hours with little effort using strong mountain updrafts to gain altitude, but other worlds may be even better suited for this mode of flight. Further terrestrial precedent includes the aerial dispersal of spider young – spiderlings crawl to the tip of a blade of grass, raise their tiny abdomens and let fly a thin silken thread, then hop aboard as a gust of wind catches the gossamer strands and whisks them away into the sky.
The idea of balloonlike living organisms is an old one both in science and science fiction. Bonnie Dalzell designed an “airship beast” for the Pick-a-Planet exhibit at the Smithsonian’s National Air and Space Museum. These creatures supposedly inhabit a world with cold winters, heavy gravity and a thick atmosphere. Twice a year the herbivorous hundred-kilogram blimps inflate their many lifting bags with metabolically generated hydrogen gas and drift to the opposite hemisphere to avoid the seasonal chill. Strong winds are an advantage, but predators are numerous and many noble aeronauts are lost during the migrations when a chance bolt of lightning strikes and ignites their flammable bodies. On Earth the Portuguese man-of-war, the chambered nautilus, and swim bladders in fishes provide precedent for a balloon lifestyle in a fluid medium.
Sail power has also been largely neglected in biology for animal locomotion. One of the few examples on this planet is Velella, a small, baggy, disk-shaped sea creature whose sail-like dorsal fin permits it to drift slowly with the wind. Another example is, surprisingly, the whale. These majestic cetaceans sometimes “stand on their heads” exposing only their giant broadleaf tails above water, catching gusts of wind and playfully “sailing” for hundreds of meters before coming up for air.
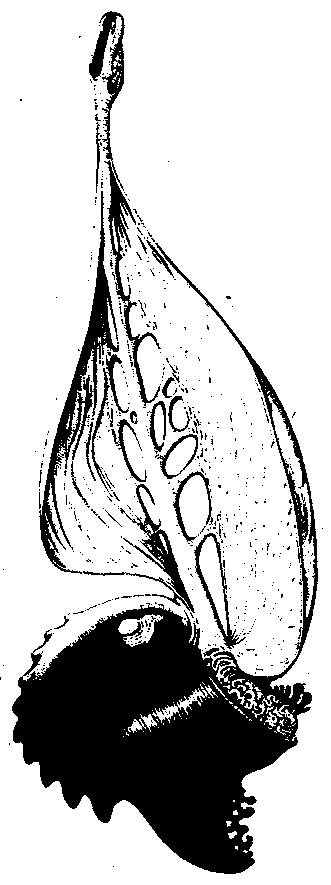
More than forty years ago Olaf Stapledon speculated on the possibility of a true biological sailboat. Let us imagine a cephalopod with a heavy concave shell living in the bays and estuaries of some alien world. Over the years the species gradually acquires the ability to float boatlike on the inverted shell as an aid in migration. These creatures drift with the shore currents, feeding on surface algae and nibbling the tops of seaweed stalks. In time the shell could become better adapted for navigation, perhaps with a streamlined undercarriage, allowing the ET to better chart its course between known patches of food and to escape its predators. Eventually it gains still more speed with a crude sail, a thin membrane growing from a shank of cartilage in the animal’s belly. With further evolution the membrane becomes retractable, even delicately manipulatable by fine muscles. At last the emergence of a brain and sensory organs strictly comparable to those of higher mollusks on Earth makes possible a kind of living clipper ship complete with masthead (forward sensors), jib, mainsail, riggings (extensible tendon), and a rudder.
Figure 4. The Sailboat Creature (courtesy of Wayne Barlowe)
Every habitable planet has millions of living species and billions of extinct ones, and there are many trillions of useful planets in the universe. This adds up to an incredible diversity of life. Christian Huygens wrote in The Celestial Worlds Discover’d (1698) that “Nature seems to court variety in her Works, and may have made them widely different from ours either in their matter or manner of Growth, in their outward Shape, or in their inward Contexture; she may have made them such as neither our Understanding nor Imagination can conceive.” Whether Huygens’s prophecy is true is something we can determine only by traveling to faraway worlds and sampling extraterrestrial ecologies at close hand. Perhaps, someday soon, we will make this epic journey.
Norman J. Berrill, Worlds Without End: A Reflection on Planets, Life and Time, Macmillan, New York, 1964.
R. McNeill Alexander, G. Goldspink, eds., Mechanics and Energetics of Animal Locomotion, John Wiley & Sons, New York, 1977.
Bonnie Dalzell, “Exotic Bestiary for Vicarious Space Voyagers,” Smithsonian 5 (October 1974):84-91.
Doris Jonas, David Jonas, Other Senses, Other Worlds, Stein and Day, New York, 1976.
Robert A. Freitas Jr., “Xenobiology,” Analog 101(30 March 1981):30-41. http://www.xenology.info/Papers/Xenobiology.htm
Olaf Stapledon, Star Maker, Methuen, 1937. Reprinted: Penguin Books, Baltimore, Maryland, 1972.
Last updated on 30 December 2008